Nitrate assimilation and nitrogen fixation are the two major pathways in the biological nitrogen cycle which provide common nitrogen sources for the synthesis and utilization of proteins and nucleic acids in all living organisms. Since the available nitrogen in the form of NH4+ in the biosphere is rather limited, a direct conversion from different inorganic forms of nitrogen to ammonia plays a crucial role, i.e. from NO3-via nitrate assimilation and from N2 by nitrogen fixation. Through nitrate assimilation, higher plants, algae, filamentous fungi, yeasts, and bacteria are able to produce more than 2 x 104 megatons of organic nitrogen per year, which is 100-times more than that produced by nitrogen fixation [1-3]. Biological conversion of nitrate to ammonium is an eight-electron reduction process with the participation of two enzymes, the reduction of nitrate to nitrite by nitrate reductase, and the reduction of nitrite to ammonium by nitrite reductase [4]. Nitrate reductase (NR) is a multi-domain enzyme comprising the prosthetic groups molybdopterin, Fe-heme, and FAD (flavin adenine dinucleotide) in a 1:1:1 stoichiometry that mediates an electron transfer from NAD(P)H to nitrate. The FAD and Mo-pterin domains function as the binding sites for NAD(P)H and NO3-, respectively [5-7], while the cytochrome b5-like heme domain facilitates the electron transfer from the FAD domain to the active-site Mo-pterin [8].
Mammalian liver cytochrome b5 is a membrane-bound protein that adds a practical difficulty in its isolation [9-11]. Recently, we have successfully cloned and expressed a cDNA encoding a water-soluble homo-tetrameric cytochrome b5-like domain (MW = 45 kDa) of Chlorella vulgaris NR in Escherichia coli [6], and have studied its electrochemical and spectroscopic properties [5,12-17]. Potentiometric titration for this recombinant truncated heme domain [5] yields a midpoint potential of +16 mV (n = 1, pH 7), which is substantially higher than that for the intact NR (-160 mV) [13] and that for the recombinant NR-heme domain coupled to a truncated Mo-pterin domain (-28 mV) [5]. The expression of a large quantity of this NR-heme domain allows the study of its structure by means of NMR techniques in order to provide more information about the correlation between its structure and physical properties. Moreover, this soluble NR-heme domain can serve as a spectroscopic model for membrane-bound cytochrome b5.
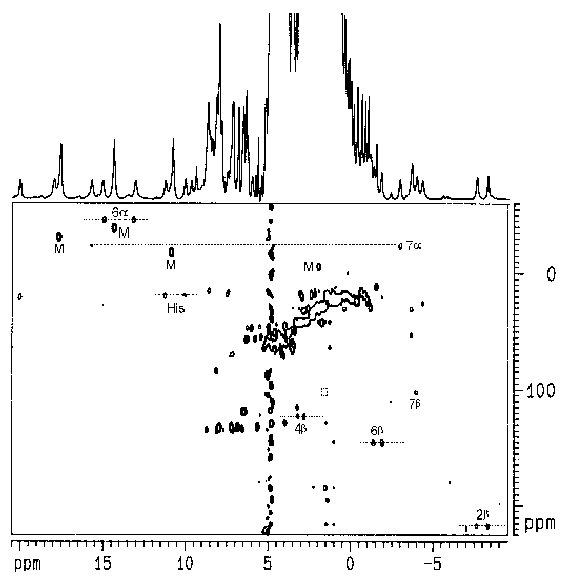
A low-spin (S = 1/2) Fe3+-heme center is known to exhibit well resolved hyperfine-shifted (i.e., isotropically shifted) 1H NMR signals attributable to the protons on the heme and in its surroundings. The study of these signals can thus reveal the structure of the heme environment. Despite its paramagnetism (which is generally not favorable for 2D NMR studies), the low-spin Fe3+-heme center in several proteins has recently been investigated by the use of various 2D NMR techniques [18-20]. Standard 2D NMR pulse sequences with appropriate adjustments of acquisition parameters can give spectra for paramagnetic molecules which provide virtually the same structural information as that obtained for diamagnetic molecules. For example: the use of coherence transfer techniques (COSY and TOCSY) allows the assignment of spin patterns, and the use of NOE techniques (NOESY) can reveal spatial arrangement of nuclei [21-23]. We have obtained a complete assignment of the isotropically shifted 1H NMR signals of the heme moiety in the truncated NR-heme domain by the use of homonuclear 1H-1H and heteronuclear 1H-13C 2D NMR techniques (see abstract). These studies represent the first steps toward a complete structural analysis of the heme-binding pocket, which is a key element for a better understanding of the unusual redox property of the heme domain in nitrate reductase.
References
[1] Dunn-Coleman, N. S., Smarelli, J. Jr. and Garrelt, R. H. (1984)
Int. Rev. Cytol. 92, 1-50.
[2] Thomsett, A. B. (1989) Microbiol. Rev. 2, 31-55.
[3] Wray, J. L. and Kinghorn, J. R. (eds.) Molecular and Genetic
Aspects of Nitrate Assimilation, Oxford Science Publications, Oxford.
[4] Solomonson, L. P. and Barber, M. J. (1990) Annu. Rev. Plant
Physiol. 41, 225-253.
[5] Cannons, A. C., Barber, M. J. and Solomonson, L. P. (1993) J.
Biol. Chem. 268, 3268-3271.
[6] Cannons, A. C., Iida, N. and Solomonson, L. P. (1991) Biochem.
J. 278, 203-209.
[7] Hyde, G. E., Crawford, N. M. and Campbell, W. H. (1991) J. Biol.
Chem. 266, No. 35, 23542-23547.
[8] Lederer, F. (1994) Biochimie 76, 674-692
[9] Meyer, C., Levin, J. M., Roussel, J.-M. and Rouze, P. (1991) J.
Biol. Chem. 266, No. 30, 20561-20566.
[10] Mathews, F. S., Levine, M. and Argos, P. (1971) Cold Spring
Harbor Symp. Quant. Biol. 36, 387-395.
[11] Mathews, F. S. and Czerwinski, E. W. (1976) The Enzymes of
Biological Membranes. Plenum Press, New York. v.4, 143.
[12] Kay, C. J. and Barber, M. J. (1986) J. Biol. Chem. 261,14125-14129.
[13] Kay, C. J., Barber, M. J. and Solomonson, L. P. (1988) Biochemistry
27, 6142-6149.
[14] Solomonson, L. P., Barber, M. J., Robbins, A. P. and Oaks, A.
(1986) J. Biol. Chem. 261, 11290-11294.
[15] Solomonson, L. P., McCreery, M. J., Kay, C. J. and Barber, M.
J. (1987) J. Biol. Chem. 262, 8934-8939.
[16] Kay, C. J., Barber, M. J., Solomonson, L. P., Kau, D., Cannons,
A. C. and Hipkin, C. R. (1990) Biochem. J. 272, 545-548.
[17] Kay, C. J., Solomonson, L. P. and Barber, M. J. (1986) J. Biol.
Chem., 261 5799-5802.
[18] Bertini, I. and Luchinat, C. (1986) NMR of Paramagnetic molecules
in biological systems. Benjamin Cummings, Menlo Park, CA.
[19] Berliner, I. and Reuben, J., Eds.(1993) NMR of Paramagnetic
Molecule, Biol. Magn. Reson. Vol 12: Plenum Press, New York, NY.
[20] La Mar, G.N., Ed. (1995). Nuclear Magnetic Resonance of Paramagnetic
Macromolecules, Kluner, Dordrecht.
[21] Ming, L.-J., Lynch, J. B., Holz, K. C. and Que, L. J. (1994) Inorg.
Chem. 33, 83-87.
[22] Ming, L.-J. and Wei, X. (1994) Inorg. Chem. 21, 4617-4618.
[23] Ming, L.-J. (1993) Magn. Reson. Chem. 31, S104-S109
{kind=link}
{kind=link}